论文《免疫系统介导的共生微生物对昆虫宿主的影响》发表在《植物保护学报》,版权归《植物保护学报》所有。本文来自网络平台,仅供参考。
先天免疫是昆虫适应复杂环境的关键,研究昆虫免疫系统有助于发现新的杀虫剂靶点,为开发新的害虫防治策略提供思路。昆虫通过模式识别受体识别环境中的病原体,激活先天免疫系统以清除病原体。昆虫的先天免疫系统主要有细胞免疫和体液免疫。Toll、免疫缺陷(immune deficiency, IMD)以及 Janus 激酶 / 信号转导和转录激活因子(Janus kinase/signal transducers and activators of transcription, JAK/STAT)是调节昆虫体液免疫的主要信号通路。然而,越来越多的研究表明共生微生物激活的免疫信号通路在行使免疫功能的同时也发挥着非免疫功能,其中胰岛素信号通路在免疫系统与宿主生长发育之间的联系中起着重要的介导作用。该文在回顾昆虫主要先天免疫信号通路和胰岛素信号通路的基础上,详细阐述了共生微生物激活的免疫信号通路对昆虫生长发育、繁殖、抗药性、化学通信、细胞更新、寿命、睡眠、能量生成途径的影响及其分子机制,并对未来的研究方向和研究手段进行了展望,以期为揭示共生微生物与昆虫宿主之间联系的生理机制提供思路。
关键词
共生微生物;免疫系统;生长发育;繁殖;抗药性;化学通信;代谢重编辑
引言
经过数十亿年的进化,生物以免疫的方式应对病原体侵染,清除异常或受损的细胞物质以维持内环境稳态 (Zhang et al., 2021)。免疫机制包括先天免疫和适应性免疫。昆虫作为地球上最繁盛的生物种群,仅仅依靠先天免疫抵抗病原体侵染。昆虫的先天免疫主要有细胞免疫和体液免疫 (Wang et al., 2023a)。Toll、免疫缺陷(immune deficiency, IMD)以及 Janus 激酶 / 信号转导和转录激活因子(Janus kinase/signal transducers and activators of transcription, JAK/STAT)是调节昆虫体液免疫的主要信号通路 (Zhang et al., 2021)。昆虫通过模式识别受体(pattern recognition receptor, PRR)识别环境中不同的病原体,激活免疫信号通路,调控免疫效应因子生成以清除病原体。
然而,越来越多的研究表明,共生微生物激活的免疫信号通路在行使免疫功能的同时也发挥着非免疫功能,对昆虫的生长发育、繁殖、化学通信、细胞更新、寿命、睡眠等生理和行为都会产生一定的影响 (Zhai et al., 2018a; Vincent et al., 2022; Darby & Lazzaro, 2023)。免疫会抑制非免疫生理活动,例如激活免疫应答会导致病原体侵染期间昆虫整体代谢速率下降、生长发育减缓、能量储备大量消耗和繁殖力下降。这些抑制效应可用生理权衡理论来解释。生理权衡是指在个体内部,一种生物学特征对另一种生物学特征的负面影响 (Schwenke et al., 2016)。该理论认为免疫应答和非免疫生理活动都需要消耗能量,而能量是有限的,当免疫系统被激活后,昆虫对其的能量供应优先于其他生理过程 (Dolezal et al., 2019)。另一方面,免疫应答也能促进昆虫某些非免疫生理活动,如细胞凋亡及增殖、化学通信和寿命等 (Zhai et al., 2018a; Vincent et al., 2022)。实际上,无论是引发抑制效应还是促进效应,昆虫在切换免疫应答和非免疫生理活动时都依赖于免疫系统与代谢信号通路间的关联来进行细胞内或细胞间,甚至是组织间的信息传递,以实现对整体代谢的调控 (Dolezal et al., 2019)。
虽然关于昆虫免疫系统的研究颇多,但对免疫信号通路发挥的非免疫作用的研究却相对滞后,相关生理机制的研究更为缺乏。例如,基于昆虫免疫细胞激活所引起的自身代谢重编辑以及全身代谢转变的研究相对于哺乳动物还有些滞后 (Dolezal et al., 2019);目前尚未明确免疫激活影响昆虫卵子生成的生理机制 (Schwenke et al., 2016)。进一步研究免疫信号通路的非免疫效应,不仅有助于深入理解免疫系统在生物体内的多效性,还有助于发现新的杀虫剂靶点,为开发新的害虫防治策略提供思路。例如,明确昆虫免疫系统对苏云金芽胞杆菌 Bacillus thuringiensis (Bt) 所产杀虫蛋白的抗性的影响有助于发现新抗性机制,从而可以开发高效的害虫综合防治技术 (廖文宇等,2022)。
1 昆虫主要免疫信号和胰岛素信号通路
1.1 Toll 免疫信号通路
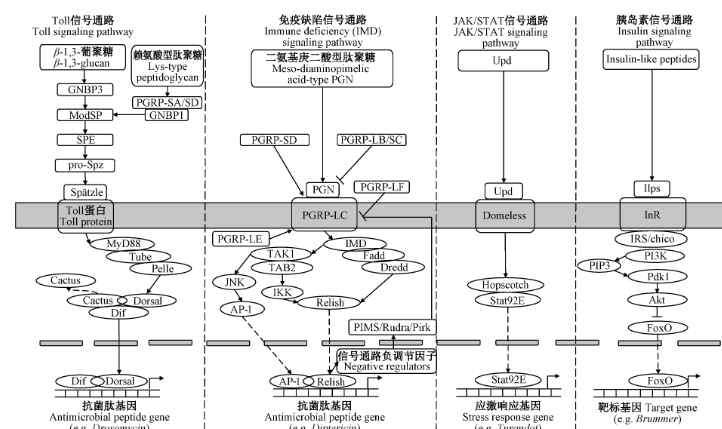
革兰氏阳性细菌产生的赖氨酸型肽聚糖(peptidoglycan, PGN)和真菌产生的 β-1,3 - 葡聚糖分别与血淋巴中的肽聚糖识别蛋白(peptidoglycan recognition protein, PGRP)和革兰氏阴性菌结合蛋白结合后 (Jang et al., 2006),进一步激活丝氨酸蛋白酶的级联反应,促使 Spӓtzle 加工酶切割胞外无活性的 Spӓtzle 前体并将其活化,活化的 Spӓtzle 与 Toll 受体结合形成 Toll 二聚体。被激活的 Toll 受体作用于胞浆蛋白髓样分化因子 88(myeloid differentiation factor 88, MyD88)/Tube(tandem ubiquitin-binding entity)/Pelle 三聚体,使得 Pelle 被磷酸化并从三聚体中分离出来,将信号传递给下游的核抑制因子 Cactus。Cactus 能阻止核转录因子 κB(nuclear factor-κB, NF-κB)或 Relish 蛋白,如转录因子 Dorsal 及其相关免疫因子(dorsal-related immunity factor, Dif)从细胞质进入细胞核内。Cactus 被泛素化和磷酸化后,Dorsal 或 Dif 从 Cactus-Dorsal 或 Cactus-Dif 复合物中释放出来并转移到细胞核中,与靶基因启动子上的 κB-DNA 元件结合进而启动抗菌肽的表达 (Zhang et al., 2021)(图 1)。
1.2 IMD 免疫信号通路
革兰氏阴性细菌和芽胞杆菌产生的二氨基庚二酸型肽聚糖与 PRR(如 PGRP-LC)结合后可激活 IMD 信号通路,进而激活 IMD 信号通路的 2 个下游分支 Relish 分支和 c-Jun 氨基末端激酶(c-Jun N-terminal kinase, JNK)分支 (胡启豪等,2019)。活化的 Relish 转入细胞核内,启动免疫相关基因的转录。PGRP-SD 和 PGRP-LE 能促进 PGN 与 PGRP-LC 的结合,增强 PGN 介导的 IMD 信号激活 (Myllymäki et al., 2014; Iatsenko et al., 2016)。
IMD 信号通路在转化生长因子 β 激活激酶 1(transforming growth factor β-activated kinase 1, TAK1)/TAK1 结合蛋白 2(TAK1-binding protein 2, TAB2)处分出了 JNK 分支。JNK 属于丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)家族成员。TAK1 是一种丝裂原活化蛋白激酶激酶激酶(mitogen activated protein kinase kinase kinase, MAP3K),TAK1 通过激活 MAPK 激酶 4(MAPK kinase 4, MKK4)及 MAPK 激酶 7(MAPK kinase 7, MKK7)而激活 JNK,继而激活激活蛋白 - 1(activating protein-1, AP-1),AP-1 进入细胞核激活应激基因的转录 (廖文宇等,2022)(图 1)。
IMD 信号通路的过度激活可能是有害的,昆虫为了保证自身正常的生长发育和繁殖进化出了相应的负调控机制。例如,在离体培养的细胞和体内所有免疫组织(血淋巴、脂肪体、成虫中肠)中,激活的 MAPK 信号通路可以诱导与 PGRP-LC 相互作用的 IMD 信号通路负调节因子 PIMS(PGRP-LC-interacting inhibitors of IMD signaling)/Rudra/Pirk 的表达 (Ragab et al., 2011)。PIMS 可阻止 PGRP-LC 迁移到细胞表面,减少细胞表面的 PRR 数量,从而抑制 IMD 信号通路的激活。具有酰胺酶活性的 IMD 信号通路负调控因子 PGRP-LB 和 PGRP-SC 可以通过降解 PGN 来抑制 IMD 信号通路 (Wang et al., 2023a)。不具有酰胺酶活性的 IMD 信号通路负调控因子 PGRP-LF 竞争性地结合 PGRP-LC,从而阻断其对 PGN 的识别 (Myllymäki et al., 2014)(图 1)。
1.3 JAK/STAT 免疫信号通路
JAK-STAT 免疫信号通路主要参与昆虫的抗病毒免疫应答,该途径不是由病毒配体直接引起的,而是由病毒侵染过程中昆虫细胞释放的细胞因子 Upd(unpaired)所诱导。病毒核酸被宿主细胞的 PRR 识别后,活化 NF-κB、干扰素调节因子以及其他转录因子,最终导致炎症因子、干扰素等细胞因子的表达 (Takeuchi & Akira, 2010)。被释放到细胞外的细胞因子与跨膜受体 Domeless 结合后,与受体蛋白结合的酪氨酸激酶 JAK 磷酸化 Hopscotch,激活后的 Hopscotch 继而磷酸化转录因子 Stat92E 并形成同源二聚体转运到细胞核中,调节下游效应基因的表达 (Cai et al., 2021)。细胞因子介导的 JAK/STAT 信号通路功能保守,其主要在细胞存活、增殖、更新、分裂分化、凋亡和免疫调节中发挥着不可或缺的作用 (Zhang et al., 2021)(图 1)。
1.4 胰岛素信号通路
胰岛素信号(insulin/insulin-like signaling, IIS)通路可通过调节昆虫体内的糖代谢、蛋白质代谢及脂质代谢来调控其生长发育和繁殖衰老等 (彭竹清和郝友进,2019)。关于昆虫 IIS 通路的研究集中在磷脂酰肌醇 - 3 - 激酶(phosphatidylinositol 3-kinase, PI3K)/ 蛋白激酶 B(protein kinase B, PKB,又称 Akt)信号途径。细胞外的胰岛素与位于细胞膜上的胰岛素受体(insulin receptor, InR)相结合,InR 被磷酸化,进而将 IIS 传递到细胞内部。IIS 在进入细胞内部后主要经由 PI3K/Akt 信号途径继续向下传递 (Yan et al., 2022)。磷酸化的 InR 激活胰岛素受体底物(insulin receptor substrate, IRS),使其与 PI3K 结合后产生磷脂酰肌醇三磷酸(phosphatidyl inositol triphosphate, PIP3),并将磷酸肌醇依赖性蛋白激酶 1(phosphoinositide-dependent protein kinase 1, Pdk1)和 Akt 招募到细胞膜上,Pdk1 在 Thr342 位点上磷酸化 Akt (Roth et al., 2018)。磷酸化的 Akt 进入到细胞质中并通过调控叉头转录因子 O 家族(fork head transcription factor O, FoxO)、糖原合成酶激酶 - 3 等下游基因的活性来行使调节功能 (Salasc et al., 2016)(图 1)。
注:箭头表示诱导效应,钉子线表示抑制效应,虚线表示转录因子跨膜。JAK/STAT:Janus 激酶 / 信号转导和转录激活因子;GNBP:革兰氏阴性菌结合蛋白;ModSP:丝氨酸蛋白酶;SPE:Spӓtzle 加工酶;pro-Spz:Spӓtzle 前体;Spätzle:胞外细胞因子;PGRP:肽聚糖识别蛋白(SA、SD、LB、SC、LF、LC、LE 均为该家族成员);MyD88、Tube、Pelle:均为胞浆蛋白;Dorsal、Dif:一种转录因子;Cactus:一种核抑制因子;Drosomycin:抗真菌肽基因;PGN:肽聚糖;TAK1:转化生长因子 β 激活激酶 1;TAB2:TAK1 结合蛋白 2;IKK:抑制核转录因子 κB 激酶;JNK:c-Jun 氨基末端激酶;AP-1:激活蛋白 - 1;Relish:一种转录因子;Fadd:Fas 相关死亡结构域蛋白;Dredd:死亡相关 ced-3/nedd2 样蛋白;PIMS/Rudra/Pirk:作用于 PGRP-LC 的 IMD 信号通路负调控因子;Diptericin:抗细菌肽基因;Upd:一种细胞因子;Domeless:DNA 黏连转录蛋白;Hopscotch:一种类 JAK 的酪氨酸激酶;Stat92E:一种转录因子;Turandot:应激响应基因;Ilps:胰岛素样肽;InR:胰岛素受体;IRS/chico:胰岛素受体底物;PI3K:磷脂酰肌醇 3 - 激酶;PIP3:磷脂酰肌醇三磷酸;Pdk1:磷酸肌醇依赖性蛋白激酶 1;Akt:蛋白激酶 B;FoxO:叉头转录因子 O 家族;Brummer:脂肪酶基因。
2 免疫激活对昆虫的影响
2.1 免疫激活对昆虫生长发育的影响
昆虫的生长发育和免疫之间存在生理权衡。病原体侵染或者基因操控激活黑腹果蝇 Drosophila melanogaster 免疫系统会导致其生长受到抑制,且能量储备减少。例如,果蝇慢性感染革兰氏阳性菌粪肠球菌 Enterococcus faecalis 会抑制其幼虫生长 (Suzawa et al., 2019);在果蝇 R4/IMDCA 突变品系幼虫的脂肪体中组成性激活 IMD 和 Toll 受体后,幼虫发育历期延长,总甘油三酯水平、脂肪体中脂质储存量和体重下降 (DiAngelo et al., 2009; Davoodi et al., 2019)。进一步的转录组研究结果证实,病原体侵染会影响碳水化合物和脂质代谢相关基因的表达 (Caragata et al., 2017),并且感染海分枝杆菌 Mycobacterium marinum 的果蝇成虫 Akt 磷酸化水平整体降低 (Dionne et al., 2006)。由于 IIS 是调节碳水化合物和脂质代谢的关键通路,对昆虫正常的生长发育至关重要,因此推测病原体侵染诱导的代谢变化可能是由昆虫免疫系统驱动的 IIS 活性变化引起的 (Darby & Lazzaro, 2023)(图 2)。
Toll 信号通路可以通过抑制 PI3K 下游 Akt 的磷酸化来抑制昆虫的生长和甘油三酯存储。如 DiAngelo et al. (2009) 研究发现在黑腹果蝇 R4/IMDCA 突变品系幼虫的脂肪体中组成性激活 Toll 信号通路后,幼虫提取物中 Akt 磷酸化水平降低;而在 MyD88 功能缺失的果蝇突变体中,Akt 磷酸化的抑制被解除。这从正反 2 个角度证实了 Toll 信号通路与 Akt 间的联系。在 PI3K 被组成性激活的果蝇幼虫脂肪体细胞中,Toll10b(Toll 样受体)的表达可以阻断细胞生长,但不影响 PIP3 的产生,表明 Toll 信号抑制 IRS/chico 和 PI3K 下游的 IIS;此外,激活的 Toll 信号通路抑制 Pdk1 在 Thr342 位点上磷酸化 Akt,从而导致果蝇生长减缓和甘油三酯储存量减少,并且 T342D 突变型 Akt(Akt 的第 342 位氨基酸突变为天冬氨酸,此突变使 Akt 处于激活状态)组成性表达可以恢复果蝇的生长和甘油三酯储存 (Roth et al., 2018)(图 2)。
除了干扰 Akt 磷酸化外,Toll 信号通路还可以通过影响胰岛素样肽(insulin-like peptide, Ilp)的合成水平来调控 IIS 通路。使用基因操控激活果蝇 Toll 信号通路后,果蝇胰岛素样肽 6(Drosophila insulin-like peptide 6, Dilp6)的合成水平降低,生长发育受到抑制 (Okamoto et al., 2009)。黑腹果蝇幼虫感染粪肠球菌后激活了 Toll 信号通路并显著降低了 Dilp6 的整体水平,在脂肪体中过表达 Dif 可降低 Dilp6 的 mRNA 转录水平并抑制幼虫生长发育,粪肠球菌侵染诱导的 Dilp6 降低和生长抑制可以通过 RNA 干扰技术敲低 Dif 来恢复 (Suzawa et al., 2019)。然而,Dif 是否直接调控 Dilp6 的表达,或者 IIS 是否受到 Dif 直接调控的蛋白的二次调控,还需要进一步的研究来验证 (Darby & Lazzaro, 2023)(图 2)。
由脂肪体中 Toll 信号驱动的昆虫生长抑制机制可能还包括激活的 Toll 信号通路诱发的外周胰岛素抵抗。胰岛素抵抗是指胰岛素作用的靶器官对胰岛素作用的敏感性下降。虽然激活的 Toll 信号通路可以通过降低脂肪体中 Dilp6 来减弱 IIS,从而抑制果蝇的生长,但这不会改变血淋巴中 Dilp2(大脑中胰岛素分泌细胞分泌的一种 InR 配体)的水平,在果蝇幼虫脂肪体中组成性激活 Toll10b 并未影响血淋巴中的 Dilp2 水平 (Brogiolo et al., 2001)。小鼠 Mus musculus 脂肪组织中 IIS 的丧失会导致其肝脏和肌肉产生胰岛素抵抗 (Shearin et al., 2016; Softic et al., 2016)。推测 Toll 信号通路下调脂肪体胰岛素合成能力后,会通过类似哺乳动物胰岛素抵抗的机制引发昆虫整体生长发育减缓 (Roth et al., 2018)。
除 Toll 信号通路外,IMD 信号通路也能介导免疫应答进而对昆虫的生长发育产生影响。尽管果蝇感染大肠埃希氏菌 Escherichia coli 激活的 IMD 和 Toll 信号通路降低了 Akt 的磷酸化,但果蝇 Toll 信号通路突变体感染大肠埃希氏菌时,对 Akt 磷酸化的抑制被解除。这表明当 Toll 和 IMD 信号通路同时被激活时,Toll 信号通路会抑制 IIS,而 IMD 信号通路则不会。此外,在果蝇脂肪体中 Relish 的组成性表达对 Akt 磷酸化水平以及甘油三酯含量无显著影响 (DiAngelo et al., 2009),但是果蝇脂肪体中 IMD 的组成性表达显著降低了全身 Akt 和 S6K(ribosomal S6 kinase)的磷酸化,幼虫生长发育延缓,甘油三酯含量降低 (Davoodi et al., 2019)。以上相反的研究结论可能归因于组成性激活 IMD 信号通路时使用了不同的果蝇基因型。推测 IMD 信号通路可能存在分支,主通路能激活 Relish,而分支通路可以通过关联其他途径来调控 IIS (Darby & Lazzaro, 2023)。此外,尽管在果蝇脂肪体中敲除 Relish 对 Akt 磷酸化、甘油三酯水平或葡萄糖含量没有影响,但敲除中肠上皮的肠内分泌细胞中的 Relish 会抑制 Dilp3 转录,增加细胞内的脂质含量,并减少 Akt 的磷酸化 (Kamareddine et al., 2018),这说明 IMD 信号通路对 IIS 的调控作用具有组织特异性(图 2)。
昆虫的 FoxO 可以对饥饿、极端温湿度、缺氧和病原体侵染等逆境做出应激反应,并在这些刺激下调节脂解和自噬等过程。即使在没有病原体侵染的情况下,饥饿也可诱导黑腹果蝇 FoxO 依赖性抗菌肽的表达,并且这些抗菌肽的表达不依赖于 Toll 和 IMD 信号通路 (Darby & Lazzaro, 2023)。FoxO 还可以激活脂肪酶基因 bmm(brummer)的转录,促进脂肪分解 (Kang et al., 2017)。在饥饿条件下,野生型果蝇的甘油三酯含量并未降低,而 Relish 缺陷基因型果蝇的甘油三酯含量显著降低,这是由 FoxO 依赖性的 bmm 转录水平上调引起的,说明 IMD 信号通路能通过调控 FoxO 活性来影响脂质代谢。进一步的研究发现,Relish 可以与 Bmm 结合,从而抑制依赖 FoxO 的脂质分解,以防止饥饿期间昆虫脂质的耗竭 (Darby & Lazzaro, 2023)。然而 Relish-FoxO 拮抗是否介导了病原体侵染期间昆虫脂质代谢的变化 (Dionne et al., 2006; Chambers et al., 2012)(图 2)尚需进一步研究验证。
注:箭头表示已知的激活效应,钉子线表示已知的抑制效应,虚线表示已知的免疫对胰岛素信号的抑制作用,但尚未明确具体机制。
2.2 免疫激活对昆虫繁殖的影响
繁殖与免疫之间也存在着生理权衡,免疫激活会抑制昆虫的繁殖力(图 3)。例如,注射大肠埃希氏菌或球孢白僵菌 Beauveria bassiana 激活黑腹果蝇雌成虫的免疫应答后,其繁殖力下降至原来的 45%(Bashir-Tanoli & Tinsley, 2014);使用含雷氏普罗威登斯菌 Providencia rettgeri 的饲料饲喂黑腹果蝇雌成虫激活其免疫应答后,其繁殖力下降 (Howick & Lazzaro, 2014);利用不同剂量的粗脂多糖激活黑腹果蝇雌成虫的免疫应答后,其繁殖力随着粗脂多糖处理剂量的增加而下降 (Nystrand & Dowling, 2014)。另外,还可以利用 IIS 活性来判定昆虫雌成虫是否有足够的营养储备生成卵子 (Schwenke et al., 2016)。高活性的 IIS 会促进昆虫生成卵子,而 IIS 活性降低时,营养储备降低,卵子生成减少或停止 (Parthasarathy & Palli, 2011)。推测病原体侵染可能通过抑制 IIS 进而抑制昆虫卵子生成 (Schwenke et al., 2016),但仍需要严谨的试验设计来验证这种调控机制。
此外,激活的 IMD 信号通路还可能通过影响神经系统来调节昆虫的产卵行为(图 3)。昆虫大脑中的神经元通过感知 PGN 来激活 IMD 信号通路,进而抑制雌成虫的产卵行为;此外昆虫大脑中还存在可以表达内源性 PGN 降解酶 PGRP-LB 的章鱼胺能神经元 (Masuzzo et al., 2019)。推测这些大脑神经元中表达的胞外负调控因子 PGRP-LB 通过调控 IMD 信号通路来调控昆虫的产卵行为 (Zhai et al., 2018a)。但是目前尚未揭示昆虫大脑中 IMD 信号调节产卵行为的机制以及血淋巴中细菌源 PGN 转移至大脑神经元的途径。小鼠肠道中的细菌源 PGN 进入血淋巴后会穿过血脑屏障到达大脑神经元,而一些神经元选择性表达 PGN 受体来感知 PGN,在昆虫中可能也具有类似的机制 (Arentsen et al., 2017),但还需要进一步研究验证。
2.3 免疫激活对昆虫抗药性的影响
IMD 信号通路可能通过影响 MAPK 信号通路来参与调节昆虫的 Bt 抗性。MAPK 信号通路是调节小菜蛾 Plutella xylostella 碱性磷酸酶(alkaline phosphatase, ALP)和三磷酸腺苷(adenosine triphosphate, ATP)结合盒(ATP-binding cassette, ABC)转运蛋白基因表达的总开关 (Guo et al., 2015)。MAPK 可介导信号从细胞表面到细胞核的转导,能作用于多种生理过程 (Davis, 1994)。ALP 和 ABCC2 是 Bt 杀虫蛋白 Cry 的受体。IMD 信号通路中的 TAK1 可以通过不同路径激活下游的 JNK 或 p38 等 MAPK,进而激活相关的转录因子,这些转录因子不仅激活应激响应基因或免疫相关基因的表达,也可能直接或者间接参与调节 ALP 和 ABCC 基因的表达,进而影响昆虫的 Bt 抗性 (廖文宇等,2022),但还需要进一步研究验证(图 3)。
昆虫肠道中双氧化酶(dual oxidase, DUOX)依赖性活性氧的产生主要由 2 个信号通路调节,一个是 DUOX 活性信号通路,该通路激活钙黏蛋白 99C(cadherin 99C, Cad99C)/ 磷脂酶 Cβ(phospholipase Cβ, PLCβ)/ 蛋白激酶 C(protein kinase C, PKC)信号,随后诱导细胞内 Ca²⁺释放,增加 DUOX 的活性;另一种是 DUOX 表达信号通路,该通路通过 MAP3K1/IMD-MKK3-p38 MAPK - 激活转录因子 2(activating transcription factor 2, Atf2)信号调节 DUOX 的表达量 (Wang et al., 2023a)。Wang et al. (2023b) 研究发现,赤拟谷盗 Tribolium castaneum 在回接肠球菌 Enterococcus sp. 后,体内 IMD 和 DUOX 的表达量上调,同时体内氧化压力增大,推测免疫系统介导的氧化压力相应增大,进而降低赤拟谷盗对磷化氢的耐受性(图 3)。
2.4 免疫激活对昆虫化学通信的影响
免疫激活会促进昆虫信息素的合成(图 3)。例如,感染棉铃虫裸病毒(Helicoverpa zea nudivirus 2, Hz-2V)的美洲棉铃虫 Helicoverpa zea 雌成虫的性信息素产量增加,其对雄成虫的引诱数量是未感染 Hz-2V 雌成虫引诱数量的 2 倍 (Burand et al., 2005);感染肠道微孢子虫 Nosema sp. 的意大利蜜蜂 Apis mellifera 的信息素释放量较未感染者增加 (Dussaubat et al., 2010)。进一步研究发现,嗜昆虫假单胞菌 Pseudomonas entomophila 通过激活 IMD 信号通路和 IIS 来改变黑腹果蝇的脂质代谢,从而增加脂肪酸衍生的月桂酸甲酯、肉豆蔻酸甲酯和棕榈酸甲酯等聚集信息素组分的释放量来吸引健康的黑腹果蝇,导致健康黑腹果蝇被感染进而增强了病原体的传播 (Keesey et al., 2017),这证实了 IMD-IIS 信号通路关联调控的脂质代谢在昆虫信息素合成中的作用。
免疫激活还会增强昆虫对病原菌的忌避行为(图 3)。当面对分别含有轻度致病性欧文氏菌 Erwinia carotovora 和高度致病性嗜昆虫假单胞菌的食物时,果蝇幼虫可以根据病原菌的气味选择取食含嗜昆虫假单胞菌的食物,而不是根据致病性的强弱选择含欧文氏菌的食物;但经过几分钟或几小时后,果蝇幼虫不再取食含嗜昆虫假单胞菌的食物 (Kobler et al., 2020)。在昆虫中,蘑菇体参与嗅觉联系学习并调节先天的气味选择行为 (Owald & Waddell, 2015)。蘑菇体功能缺失的果蝇不再偏好含有某种病原菌的食物 (Kobler et al., 2020)。进一步研究发现,具蘑菇体调节功能的昆虫对微生物的适应行为需要章鱼胺能神经元的参与 (Masuzzo et al., 2019)。如果章鱼胺能神经元中的 PGRP-LC/LE 表达水平下调,果蝇则更偏好含病原菌的蔗糖,并且果蝇 PGRP-LC/LE 缺失突变体对上述 2 种病原菌没有选择偏好 (Kobler et al., 2020)。这些结果表明 IMD 信号通路参与将病原体摄入信号传递到神经系统的过程。病原菌激活果蝇的免疫应答,诱导章鱼胺能神经元表达 PGRP-LC/LE 以识别病原菌 (Harris et al., 2015),并将信息传递给蘑菇体,而蘑菇体持续调控摄食病原菌行为的机制可能类似于短期嗅觉联系学习 (Kobler et al., 2020)。在昆虫中 PGN 可被释放到血淋巴并转送到其他器官 (Basset et al., 2000)。同样,在哺乳动物中 PGN 可以被转运到大脑,被 PGRP 识别进而调节其社会行为 (Arentsen et al., 2017)。因此,推测肠道摄入的细菌源 PGN 从消化系统进入果蝇大脑,进而激活神经系统中的 PGRP-LC 信号 (Kobler et al., 2020)(图 3)。
但是,有研究认为果蝇对病原菌的忌避行为可能是因为病原菌侵染调节了果蝇的嗅觉识别能力。果蝇取食被胡萝卜软腐欧文氏菌胡萝卜亚种 Erwinia carotovora subsp. carotovora 污染的食物,其肠道会产生细胞因子来激活触角叶中围鞘胶质细胞的 JAK/STAT 信号通路,从而诱导单羧酸转运蛋白和载脂胶质蛋白 Lazarillo 的表达 (Cai et al., 2021)。单羧酸转运蛋白可以实现乳酸在神经元和胶质细胞间的转运,进而促进神经元中脂质生成和胶质细胞内脂滴积累,而 Lazarillo 能促进脂质从神经元到胶质细胞的转运,进而影响触角叶中胶质细胞与神经元的代谢耦合,增强了果蝇的嗅觉识别能力,引发其对病原菌污染食物的忌避行为,增加了存活率 (Cai et al., 2021)(图 3)。
2.5 免疫激活对昆虫细胞更新的影响
免疫激活会促进昆虫细胞的凋亡和增殖。在衰老的昆虫肠道中,细菌总载量增加,并且肠道微生物群落中的革兰氏阴性醋酸杆菌属 Acetobacteria 载量急剧增加;而为了控制肠道菌的载量,IMD 信号通路被过度激活 (Buchon et al., 2009a; Guo et al., 2014)。通过基因操控过表达 IMD 激活果蝇 IMD 信号通路后,脂肪体细胞的凋亡速度加快 (Georgel et al., 2001)。细菌侵染激活的 IMD 通路会诱导果蝇中肠的肠壁细胞脱落到肠腔,而在 PGRP-LE/IMD/ 死亡相关 ced-3/nedd2 样蛋白(death related ced-3/nedd2-like protein, Dredd)/Relish 功能缺失的果蝇突变体中,其肠壁细胞脱落被抑制 (Zhai et al., 2018b)。此外,高活性的 IMD 还可以加速果蝇肠壁细胞的更新并促进肠道干细胞的补偿性增殖 (Buchon et al., 2009b; Jiang et al., 2009)。激活的 IMD 信号通路还会促进果蝇肠道蠕动,有助于将入侵细菌排出体外,提高果蝇的免疫力 (Hori et al., 2018)。此外,在非免疫组织中,对免疫通路进行适当的负调控有助于正常的细胞更新。在 PGRP-LF 功能缺失的果蝇突变体中,其外胚层细胞凋亡被抑制,导致其在化蛹过程中不能正常形成前胸背板和生殖器 (Tavignot et al., 2017)(图 3)。
除 IMD 信号通路外,JAK/STAT 信号通路也会促进昆虫细胞更新。胡萝卜软腐欧文氏菌胡萝卜亚种侵染果蝇后会导致其中肠肠壁细胞受损,进而产生炎症细胞因子 Upd 2 和 Upd 3,这些细胞因子进一步激活肠道干细胞中的 JAK-STAT 信号通路,促进果蝇肠道干细胞增殖和肠壁细胞再生 (Buchon et al., 2009b; Jiang et al., 2009)(图 3)。
2.6 免疫激活对昆虫寿命的影响
激活的 IMD 信号通路可以延长果蝇寿命。PGRP-LC 或 Relish 突变的果蝇品系寿命缩短 (Buchon et al., 2009a)。此外,PGRP-SD 通过拮抗 IMD 信号通路的胞外负调控因子 PGRP-LB 来增加 IMD 信号的强度 (Iatsenko et al., 2016)。PGRP-SD 过表达可以防止果蝇肠道微生物菌群失调,从而延长其寿命,而在 PGRP-SDsk1 突变的果蝇肠道中植物乳植杆菌 Lactiplantibacillus plantarum 的载量增加,增大了昆虫体内的氧化压力,使其寿命缩短 (Iatsenko et al., 2018)。但是,过度激活 IMD 信号通路会缩短果蝇的寿命。在衰老的果蝇肠道中,通过激活转录因子 FoxO 可以抑制 IMD 信号通路的负调节因子 PGRP-SC2 的表达,从而引起 Relish/NF-κB 活性失调,导致干细胞过度增殖和肠壁细胞异常增生,而恢复 PGRP-SC2 在肠壁细胞中的表达能促进组织稳态,进而延长果蝇寿命 (Guo et al., 2014)(图 3)。
2.7 免疫激活对昆虫睡眠的影响
免疫激活会影响果蝇睡眠,如系统感染肺炎链球菌 Streptococcus pneumoniae 后,黑腹果蝇成虫的睡眠时间缩短,昼夜节律紊乱 (Shirasu-Hiza et al., 2007)。使用 Geneswitch 技术上调果蝇成虫的 PGRP-LCa 激活 IMD 信号通路后,果蝇睡眠时间缩短 (Mallon et al., 2014)。相反地,系统感染大肠埃希氏菌激活免疫应答后,黑腹果蝇成虫睡眠时间延长,延长的程度受 Relish 表达量的影响,而黑腹果蝇 Relish 突变体感染大肠埃希氏菌后,其睡眠时间没有增加 (Kuo et al., 2010)。目前对感染过程中果蝇睡眠是被诱导还是被抑制的结论仍存在争议,不同的研究结论可能是因为果蝇感染病原菌的时段不同 (Lee & Edery, 2008),也可能是所用病原菌及其激活的免疫因子不同 (Vincent et al., 2022)(图 3)。
2.8 细胞免疫激活引起的代谢重编辑
昆虫的免疫系统包括体液免疫和细胞免疫,无论何种免疫都需要高效的能量供给。为满足自身的能量需求,与哺乳动物巨噬细胞中的 Warburg 效应类似,昆虫的免疫细胞会进行代谢重编辑。Warburg 效应是指即使在足够的氧气供应下,免疫细胞更倾向于通过糖酵解产生乳酸生成 ATP,而不是通过氧化磷酸化(oxidative phosphorylation, OXPHOS)产能 (Dolezal et al., 2019)。
免疫激活会引起免疫细胞自身的代谢重编辑(图 3)。静止状态的免疫细胞利用最有效的糖酵解 - 三羧酸循环 - OXPHOS 途径产生 ATP(理论上每分子葡萄糖产生 38 个 ATP),以满足基础能量需求 (Palsson-McDermott & O’Neill, 2013)。虽然这种能量生成方式非常有效,但是产生 ATP 的速度较慢,不能满足免疫应答期间免疫细胞对能量的快速需求,因此,免疫细胞将能量生成途径切换为乳酸生成途径,该途径的产能效率虽然不高(每分子葡萄糖只产生 2 分子 ATP),但其产能速率要比以糖酵解 - 三羧酸循环 - OXPHOS 途径快得多 (Dolezal et al., 2019)。
在哺乳动物中,免疫激活的 NF-κB 信号介导了低氧诱导因子 - 1α(hypoxia inducible factor-1α, HIF-1α)诱导的免疫细胞代谢重编辑。HIF-1α 负责启动缺氧引起的细胞代谢重编辑 (Wenger et al., 2005)。HIF-1α 的水平受氧浓度的严格调控,在有氧条件下,HIF-1α 的特定脯氨酸残基被脯氨酸羟化酶(prolyl hydroxylase dehydrogenase, PHD)羟基化,这是 HIF-1α 被降解的标志。在有氧条件下,激活的哺乳动物 Toll 样受体和 NF-κB 信号通过铁蛋白介导的信号途径抑制 PHD,使 HIF-1α 的表达量增加并趋于稳定化 (Siegert et al., 2015)。高水平的 HIF-1α 会促进机体摄入葡萄糖,增强免疫细胞的糖酵解代谢途径和戊糖磷酸化途径,同时抑制三羧酸循环,其中乳酸脱氢酶(促进丙酮酸转化为乳酸)、乳酸转运体、一氧化氮合酶、一氧化氮(对线粒体电子传递链产生亚硝化作用,大量减少 OXPHOS)以及丙酮酸脱氢酶激酶(抑制丙酮酸脱氢酶,阻碍丙酮酸进入三羧酸循环)的表达量提高 (Dolezal et al., 2019)。
在有氧条件下,通过遗传或感染激活 Toll 和 IMD 信号通路诱导的 NF-κB 信号均能诱导果蝇免疫细胞中 HIF-1α 的表达。如在褪色沙雷氏菌 Serratia marcescens 侵染果蝇期间,IMD/IκB 激酶(inhibitor of NF-κB kinase, IKK)/Relish 能诱导 HIF-1α 的表达,敲除 HIF-1α 导致果蝇感染期间的死亡率增加 (Bandarra et al., 2015)。通过基因操控过表达果蝇 Toll 信号通路的 NF-κB 因子 Dorsal 或 Cactus 编码基因均会诱导 HIF-1α 的表达 (van Uden et al., 2011)。烟芽夜蛾 Heliothis virescens 幼虫的免疫系统被激活后,其血细胞转录组分析结果表明糖酵解基因和乳酸脱氢酶基因的表达量增加 (Shelby & Popham, 2012)。在果蝇幼虫被寄生蜂寄生期间,一种免疫细胞 —— 层状细胞的增殖和分化与血细胞特异性糖酵解基因表达的增加有关,并伴随着葡萄糖消耗的增加和乳酸的产生 (Bajgar et al., 2015; Strasser, 2016)。虽然各种昆虫免疫细胞在激活后显著增加了葡萄糖消耗、糖酵解和乳酸产生,但是昆虫和哺乳动物的代谢差异较大,有必要进一步详细研究昆虫相关代谢的变化。
免疫激活同样会对昆虫全身的代谢产生影响(图 3)。在昆虫免疫应答期间,免疫细胞对葡萄糖的摄取会对整体代谢产生一定影响,并且影响力度取决于免疫应答的程度。例如,在寄生蜂寄生期间,果蝇幼虫激活免疫细胞需要消耗大量葡萄糖 (Bajgar et al., 2015)。这种情况下免疫激活会显著抑制寄生期间宿主的其他代谢,如寄生蜂寄生抑制了果蝇幼虫发育过程中甘油三酯和糖原的储备 (Bajgar & Dolezal, 2018),同样地,单核增生李斯特氏菌 Listeria monocytogenes 侵染也会抑制黑腹果蝇发育过程中甘油三酯和糖原的储备 (Chambers et al., 2012)。进一步研究发现,腺苷介导了免疫激活期间昆虫整体代谢的转换,腺苷以消耗能量储备和抑制整体代谢为代价,为昆虫免疫细胞的快速增殖和层状细胞的分化以及寄生卵的有效包囊提供所需能量,或为它们有效吞噬细菌提供所需能量 (Dolezal et al., 2019)。果蝇血淋巴中的血细胞识别寄生蜂产生的卵后激活专性免疫细胞,加速层状细胞的增殖和分化,层状细胞形成包囊并破坏寄生蜂的卵,被激活的层状细胞前体糖酵解和葡萄糖消耗增加,并通过释放腺苷从昆虫的其余组织争夺葡萄糖;而胞外腺苷通过腺苷受体传递信号来抑制其他组织的代谢,从而减缓幼虫的发育,并通过释放腺苷调控整体代谢,使免疫系统具有能量使用优先权 (Bajgar et al., 2015)。
注:箭头表示诱导效应,钉子线表示抑制效应,虚线表示尚未明确是诱导效应还是抑制效应。IMD:免疫缺陷;JNK:c-Jun 氨基末端激酶;DUOX:双氧化酶;JAK/STAT:Janus 激酶 / 信号转导和转录激活因子。
3 展望
越来越多的研究证实免疫信号通路是昆虫防御、生理和行为之间的协调者,但是目前尚未明确免疫信号通路转录因子下游基因的功能特征,因此,对于免疫信号通路调节除免疫外的繁殖、抗药性、化学通信、细胞更新、寿命、睡眠等其他功能的认识仍处于初级阶段。为揭示免疫信号通路调控昆虫生理和行为的机制,需要重点明确以下几个问题:一是免疫激活的 IMD 信号通路如何关联 IIS 来调控昆虫卵子生成;二是 IMD 信号通路是否通过 MAPK 信号通路调节 ALP 和 ABC 转运蛋白等 Bt 受体的表达,从而改变昆虫对 Bt 的抗性;三是免疫激活的 IMD 信号通路如何关联 IIS 来调控脂质代谢,进而影响昆虫信息素的合成;四是免疫激活的 IMD 和 Toll 信号通路是否介导了 HIF-1α 诱导的昆虫免疫细胞的代谢重编辑以及整体代谢的转变。
在研究免疫系统介导的共生微生物对昆虫的影响时,激活免疫的途径有 3 种,即经口感染、系统感染和遗传操控。经口感染是一种通过消化道摄入病原体以激活昆虫免疫系统的方法,可以模拟自然感染过程,常用于研究昆虫的肠道免疫,如经口感染欧文氏菌会导致依赖于 Relish 的中肠基因表达水平发生变化,包括免疫和代谢基因调节因子 (Buchon et al., 2009b)。系统感染是通过显微注射的方法将病原体引入昆虫体内,从而引发全身的免疫反应,如系统感染藤黄微球菌 Micrococcus luteus 的黑腹果蝇脂肪体中 Akt 磷酸化水平和甘油三酯含量降低 (DiAngelo et al., 2009)。遗传操控是通过敲除、过表达或基因编辑等基因工程技术改造生物体的基因,以激活或抑制特定的免疫反应,多用来研究特定基因在免疫系统中的功能或对其他生理功能的影响,如使用上游激活序列 / 半乳糖调节的上游启动子元件(upstream activating sequence/galactose regulated upstream promoter element, UAS-Gal4)在果蝇脂肪体中过表达 Toll 会抑制 Akt 磷酸化,降低甘油三酯含量 (Roth et al., 2018; Suzawa et al., 2019)。这 3 种免疫激活方法各有优势,选择哪种方法取决于研究的具体目标。
尽管对昆虫免疫与代谢互作的研究相对于哺乳动物免疫系统的研究有些滞后,但最近的研究发现在免疫应答期间,昆虫和哺乳动物免疫与代谢互作的机制非常相似,因此,哺乳动物在免疫细胞激活所引起的自身代谢重编辑以及全身代谢转变方面的研究为昆虫免疫细胞代谢的深入研究提供了思路。此外,代谢组学分析技术的快速发展有助于开展免疫与代谢互作相关研究 (王争艳等,2020)。其中,基质辅助激光解吸离子化质谱成像技术可以实现完整组织样品内代谢物空间分布的可视化,在细胞特异性代谢组学研究中具有较好的应用前景 (Cox et al., 2017)。目前已有许多以黑腹果蝇、家蚕 Bombyx mori 和斯氏按蚊 Anopheles stephensi 等昆虫为模型的体内感染模型 (Darby & Lazzaro, 2023),未来可以结合这些技术研究免疫细胞的代谢。与小鼠相比,昆虫相对简单,尤其是遗传可操作性优良的果蝇已经成为研究组织间信息交流的良好模型 (Droujinine & Perrimon, 2016),可用于研究免疫应答过程中整体代谢的调节和组织间的信息交流,揭示免疫和代谢之间的相互作用。
参考文献 (References)
[1] Arentsen T, Qian Y, Gkotzis S, Femenia T, Wang T, Udekwu K, Forssberg H, Diaz Heijtz R. 2017. The bacterial peptidoglycan-sensing molecule Pglyrp2 modulates brain development and behavior[J]. Molecular Psychiatry, 22(2): 257-266.
[2] Bajgar A, Dolezal T. 2018. Extracellular adenosine modulates host-pathogen interactions through regulation of systemic metabolism during immune response in Drosophila[J]. PLoS Pathogens, 14(4): e1007022.
[3] Bajgar A, Kucerova K, Jonatova L, Tomcala A, Schneedorferova I, Okrouhlik J, Dolezal T. 2015. Extracellular adenosine mediates a systemic metabolic switch during immune response[J]. PLoS Biology, 13(4): e1002135.
[4] Bandarra D, Biddlestone J, Mudie S, Müller HAJ, Rocha S. 2015. HIF1α restricts NF-κB-dependent gene expression to control innate immunity signals[J]. Disease Models & Mechanisms, 8(2): 169-181.
[5] Bashir-Tanoli S, Tinsley MC. 2014. Immune response costs are associated with changes in resource acquisition and not resource reallocation[J]. Functional Ecology, 28(4): 1011-1019.
[6] Basset A, Khush RS, Braun A, Gardan L, Boccard F, Hoffmann JA, Lemaitre B. 2000. The phytopathogenic bacteria Erwinia carotovora infects Drosophila and activates an immune response[J]. Proceedings of the National Academy of Sciences of the United States of America, 97(7): 3376-3381.
[7] Brogiolo W, Stocker H, Ikeya T, Rintelen F, Fernandez R, Hafen E. 2001. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control[J]. Current Biology, 11(4): 213-221.
[8] Buchon N, Broderick NA, Chakrabarti S, Lemaitre B. 2009a. Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila[J]. Genes & Development, 23(19): 2333-2344.
[9] Buchon N, Broderick NA, Poidevin M, Pradervand S, Lemaitre B. 2009b. Drosophila intestinal response to bacterial infection: activation of host defense and stem cell proliferation[J]. Cell Host & Microbe, 5(2): 200-211.
[10] Burand JP, Tan WJ, Kim WJ, Nojima S, Roelofs W. 2005. Infection with the insect virus Hz-2v alters mating behavior and pheromone production in female Helicoverpa zea moths[J]. Journal of Insect Science, 5: 6.
[11] Cai XT, Li HJ, Borch Jensen M, Maksoud E, Borneo J, Liang YX, Quake SR, Luo LQ, Haghighi P, Jasper H. 2021. Gut cytokines modulate olfaction through metabolic reprogramming of glia[J]. Nature, 596(7870): 97-102.
[12] Caragata EP, Pais FS, Baton LA, Silva JBL, Sorgine MHF, Moreira LA. 2017. The transcriptome of the mosquito Aedes fluviatilis (Diptera: Culicidae), and transcriptional changes associated with its native Wolbachia infection[J]. BMC Genomics, 18(1): 6.
[13] Chambers MC, Song KH, Schneider DS. 2012. Listeria monocytogenes infection causes metabolic shifts in Drosophila melanogaster[J]. PLoS ONE, 7(12): e50679.
[14] Cox JE, Thummel CS, Tennessen JM. 2017. Metabolomic studies in Drosophila[J]. Genetics, 206(3): 1169-1185.
[15] Darby AM, Lazzaro BP. 2023. Interactions between innate immunity and insulin signaling affect resistance to infection in insects[J]. Frontiers in Immunology, 14: 1276357.
[16] Davis RJ. 1994. MAPKs: new JNK expands the group[J]. Trends in Biochemical Sciences, 19(11): 470-473.
[17] Davoodi S, Galenza A, Panteluk A, Deshpande R, Ferguson M, Grewal S, Foley E. 2019. The immune deficiency pathway regulates metabolic homeostasis in Drosophila[J]. Journal of Immunology, 202(9): 2747-2759.
[18] DiAngelo JR, Bland ML, Bambina S, Cherry S, Birnbaum MJ. 2009. The immune response attenuates growth and nutrient storage in Drosophila by reducing insulin signaling[J]. Proceedings of the National Academy of Sciences of the United States of America, 106(49): 20853-20858.
[19] Dionne MS, Pham LN, Shirasu-Hiza M, Schneider DS. 2006. Akt and foxo dysregulation contribute to infection-induced wasting in Drosophila[J]. Current Biology, 16(20): 1977-1985.
[20] Dolezal T, Krejcova G, Bajgar A, Nedbalova P, Strasser P. 2019. Molecular regulations of metabolism during immune response in insects[J]. Insect Biochemistry and Molecular Biology, 109: 31-42.
[21] Droujinine IA, Perrimon N. 2016. Interorgan communication pathways in physiology: focus on Drosophila[J]. Annual Review of Genetics, 50: 539-570.
[22] Dussaubat C, Maisonnasse A, Alaux C, Tchamitchan S, Brunet JL, Plettner E, Belzunces LP, Le Conte Y. 2010. Nosema spp. infection alters pheromone production in honey bees (Apis mellifera)[J]. Journal of Chemical Ecology, 36(5): 522-525.
[23] Georgel P, Naitza S, Kappler C, Ferrandon D, Zachary D, Swimmer C, Kopczynski C, Duyk G, Reichhart JM, Hoffmann JA. 2001. Drosophila immune deficiency (IMD) is a death domain protein that activates antibacterial defense and can promote apoptosis[J]. Developmental Cell, 1(4): 503-514.
[24] Guo LL, Karpac J, Tran SL, Jasper H. 2014. PGRP-SC2 promotes gut immune homeostasis to limit commensal dysbiosis and extend lifespan[J]. Cell, 156(1/2): 109-122.
[25] Guo ZJ, Kang S, Chen DF, Wu QJ, Wang SL, Xie W, Zhu X, Baxter SW, Zhou XG, Jurat-Fuentes JL, et al. 2015. MAPK signaling pathway alters expression of midgut ALP and ABCC genes and causes resistance to Bacillus thuringiensis Cry1Ac toxin in diamondback moth[J]. PLoS Genetics, 11(4): e1005124.
[26] Harris N, Braiser DJ, Dickman DK, Fetter RD, Tong A, Davis GW. 2015. The innate immune receptor PGRP-LC controls presynaptic homeostatic plasticity[J]. Neuron, 88(6): 1157-1164.
[27] Hori A, Kurata S, Kuraishi T. 2018. Unexpected role of the IMD pathway in Drosophila gut defense against Staphylococcus aureus[J]. Biochemical and Biophysical Research Communications, 495(1): 395-400.
[28] Howick VM, Lazzaro BP. 2014. Genotype and diet shape resistance and tolerance across distinct phases of bacterial infection[J]. BMC Evolutionary Biology, 14(1): 56.
[29] 胡启豪, 卢玉珍, 余小强. 2019. 黑腹果蝇先天免疫研究进展[J]. 应用昆虫学报, 56(5): 947-956.
[30] Iatsenko I, Boquete JP, Lemaitre B. 2018. Microbiota-derived lactate activates production of reactive oxygen species by the intestinal NADPH oxidase Nox and shortens Drosophila lifespan[J]. Immunity, 49(5): 929-942.
[31] Iatsenko I, Kondo S, Mengin-Lecreulx D, Lemaitre B. 2016. PGRP-SD, an extracellular pattern-recognition receptor, enhances peptidoglycan-mediated activation of the Drosophila IMD pathway[J]. Immunity, 45(5): 1013-1023.
[32] Jang IH, Chosa N, Kim SH, Nam HJ, Lemaitre B, Ochiai M, Kambris Z, Brun S, Hashimoto C, Ashida M, et al. 2006. A spätzle-processing enzyme required for Toll signaling activation in Drosophila innate immunity[J]. Developmental Cell, 10(1): 45-55.
[33] Jiang HQ, Patel PH, Kohlmaier A, Grenley MO, McEwen DG, Edgar BA. 2009. Cytokine/Jak/Stat signaling mediates regeneration and homeostasis in the Drosophila midgut[J]. Cell, 137(7): 1343-1355.
[34] Kamareddine L, Robins WP, Berkey CD, Mekalanos JJ, Watnick PI. 2018. The Drosophila immune deficiency pathway modulates enteroendocrine function and host metabolism[J]. Cell Metabolism, 28(3): 449-462.
[35] Kang P, Chang K, Liu Y, Bouska M, Birnbaum A, Karashchuk G, Thakore R, Zheng WJ, Post S, Brent CS, et al. 2017. Drosophila Kruppel homolog 1 represses lipolysis through interaction with dFOXO[J]. Scientific Reports, 7(1): 16369.
[36] Keesey IW, Koerte S, Khallaf MA, Retzke T, Guillou A, Grosse-Wilde E, Buchon N, Knaden M, Hansson BS. 2017. Pathogenic bacteria enhance dispersal through alteration of Drosophila social communication[J]. Nature Communications, 8(1): 265.
[37] Kobler JM, Rodriguez Jimenez FJ, Petcu I, Grunwald Kadow IC. 2020. Immune receptor signaling and the mushroom body mediate post-ingestion pathogen avoidance[J]. Current Biology, 30(23): 4693-4709.
[38] Kuo TH, Pike DH, Beizaeipour Z, Williams JA. 2010. Sleep triggered by an immune response in Drosophila is regulated by the circadian clock and requires the NFκB Relish[J]. BMC Neuroscience, 11: 17.
[39] Lee JE, Edery I. 2008. Circadian regulation in the ability of Drosophila to combat pathogenic infections[J]. Current Biology, 18(3): 195-199.
[40] 廖文宇, 吕卓鸿, 张友军, 杨中侠. 2022. 害虫Bt抗性机制研究新方向: 昆虫体液免疫系统[J]. 昆虫学报, 65(11): 1547-1564.
[41] Mallon EB, Alghamdi A, Holdbrook RTK, Rosato E. 2014. Immune stimulation reduces sleep and memory ability in Drosophila melanogaster[J]. PeerJ, 2: e434.
[42] Masuzzo A, Manière G, Viallat-Lieutaud A, Avazeri É, Zugasti O, Grosjean Y, Kurz CL, Royet J. 2019. Peptidoglycan-dependent NF-κB activation in a small subset of brain octopaminergic neurons controls female oviposition[J]. eLife, 8: e50559.
[43] Myllymäki H, Valanne S, Rämet M. 2014. The Drosophila IMD signaling pathway[J]. Journal of Immunology, 192(8): 3455-3462.
[44] Nystrand M, Dowling DK. 2014. Dose-dependent effects of an immune challenge at both ultimate and proximate levels in Drosophila melanogaster[J]. Journal of Evolutionary Biology, 27(5): 876-888.
[45] Okamoto N, Yamanaka N, Yagi Y, Nishida Y, Kataoka H, O’Connor MB, Mizoguchi A. 2009. A fat body-derived IGF-like peptide regulates postfeeding growth in Drosophila[J]. Developmental Cell, 17(6): 885-891.
[46] Owald D, Waddell S. 2015. Olfactory learning skews mushroom body output pathways to steer behavioral choice in Drosophila[J]. Current Opinion in Neurobiology, 35: 178-184.
[47] Palsson-McDermott EM, O’Neill LAJ. 2013. The Warburg effect then and now: from cancer to inflammatory diseases[J]. BioEssays, 35(11): 965-973.
[48] Parthasarathy R, Palli SR. 2011. Molecular analysis of nutritional and hormonal regulation of female reproduction in the red flour beetle, Tribolium castaneum[J]. Insect Biochemistry and Molecular Biology, 41(5): 294-305.
[49] 彭竹清, 郝友进. 2019. 胰岛素信号调控昆虫滞育的研究进展[J]. 上海交通大学学报(农业科学版), 37(6): 201-206.
[50] Ragab A, Buechling T, Gesellchen V, Spirohn K, Boettcher AL, Boutros M. 2011. Drosophila Ras/MAPK signalling regulates innate immune responses in immune and intestinal stem cells[J]. The EMBO Journal, 30(6): 1123-1136.
[51] Roth SW, Bitterman MD, Birnbaum MJ, Bland ML. 2018. Innate immune signaling in Drosophila blocks insulin signaling by uncoupling PI(3,4,5)P3 production and Akt activation[J]. Cell Reports, 22(10): 2550-2556.
[52] Salasc F, Mutuel D, Debaisieux S, Perrin A, Dupressoir T, Grenet ASG, Ogliastro M. 2016. Role of the phosphatidylinositol-3-kinase/Akt/target of rapamycin pathway during ambidensovirus infection of insect cells[J]. Journal of General Virology, 97(1): 233-245.
[53] Schwenke RA, Lazzaro BP, Wolfner MF. 2016. Reproduction-immunity trade-offs in insects[J]. Annual Review of Entomology, 61: 239-256.
[54] Shearin AL, Monks BR, Seale P, Birnbaum MJ. 2016. Lack of AKT in adipocytes causes severe lipodystrophy[J]. Molecular Metabolism, 5(7): 472-479.
[55] Shelby KS, Popham HJR. 2012. RNA-Seq study of microbially induced hemocyte transcripts from larval Heliothis virescens (Lepidoptera: Noctuidae)[J]. Insects, 3(3): 743-762.
[56] Shirasu-Hiza MM, Dionne MS, Pham LN, Ayres JS, Schneider DS. 2007. Interactions between circadian rhythm and immunity in Drosophila melanogaster[J]. Current Biology, 17(10): R353-R355.
[57] Siegert I, Schödel J, Nairz M, Schatz V, Dettmer K, Dick C, Kalucka J, Franke K, Ehrenschwender M, Schley G, et al. 2015. Ferritin-mediated iron sequestration stabilizes hypoxia-inducible factor-1α upon LPS activation in the presence of ample oxygen[J]. Cell Reports, 13(10): 2048-2055.
[58] Softic S, Boucher J, Solheim MH, Fujisaka S, Haering MF, Homan EP, Winnay J, Perez-Atayde AR, Kahn CR. 2016. Lipodystrophy due to adipose tissue-specific insulin receptor knockout results in progressive NAFLD[J]. Diabetes, 65(8): 2187-2200.
[59] Strasser P. 2016. Warburg effect in lymph gland of Drosophila melanogaster upon parasitoid wasp infection[D]. Ceske Budejovice: University of South Bohemia, 1-36.
[60] Suzawa M, Muhammad NM, Joseph BS, Bland ML. 2019. The toll signaling pathway targets the insulin-like peptide Dilp6 to inhibit growth in Drosophila[J]. Cell Reports, 28(6): 1439-1446.
[61] Takeuchi O, Akira S. 2010. Pattern recognition receptors and inflammation[J]. Cell, 140(6): 805-820.
[62] Tavignot R, Chaduli D, Djitte F, Charroux B, Royet J. 2017. Inhibition of a NF-κB/Diap1 pathway by PGRP-LF is required for proper apoptosis during Drosophila development[J]. PLoS Genetics, 13(1): e1006569.
[63] van Uden P, Kenneth NS, Webster R, Müller HA, Mudie S, Rocha S. 2011. Evolutionary conserved regulation of HIF-1β by NF-κB[J]. PLoS Genetics, 7(1): e1001285.
[64] Vincent CM, Beckwith EJ, Simoes da Silva CJ, Pearson WH, Kierdorf K, Gilestro GF, Dionne MS. 2022. Infection increases activity via Toll dependent and independent mechanisms in Drosophila melanogaster[J]. PLoS Pathogens, 18(9): e1010826.
[65] 王争艳, 苗世远, 何梦婷, 王文芳, 鲁玉杰. 2020. 昆虫共生菌对宿主功能研究的方法体系[J]. 应用昆虫学报, 57(3): 744-758.
[66] Wang ZY, Yong HZ, Zhang S, Liu ZY, Zhao YR. 2023a. Colonization resistance of symbionts in their insect hosts[J]. Insects, 14(7): 594.
[67] Wang ZY, Zhang S, Liu ZY, Chang ZZ, Hu HS. 2023b. Gut bacteria promote phosphine susceptibility of Tribolium castaneum by aggravating oxidative stress and fitness costs[J]. Insects, 14(10): 815.
[68] Wenger RH, Stiehl DP, Camenisch G. 2005. Integration of oxygen signaling at the consensus HRE[J]. Science’s STKE, 2005(306): re12.
[69] Yan JM, Xue Z, Dong HN, Pang JX, Liu HW, Gong J, Xia QY, Hou Y. 2022. Nutrition regulates the expression of storage proteins in Bombyx mori via insulin-like/FoxO signaling pathway[J]. Insect Biochemistry and Molecular Biology, 149: 103847.
[70] Zhai ZZ, Huang XS, Yin YL. 2018a. Beyond immunity: the IMD pathway as a coordinator of host defense, organismal physiology and behavior[J]. Developmental & Comparative Immunology, 83: 51-59.
[71] Zhai ZZ, Boquete JP, Lemaitre B. 2018b. Cell-specific IMD-NF-κB responses enable simultaneous antibacterial immunity and intestinal epithelial cell shedding upon bacterial infection[J]. Immunity, 48(5): 897-910.
[72] Zhang W, Tettamanti G, Bassal T, Heryanto C, Eleftherianos I, Mohamed A. 2021. Regulators and signalling in insect antimicrobial innate immunity: functional molecules and cellular pathways[J]. Cellular Signalling, 83: 110003.



