为探明喀斯特地区次生林优势种群竞争关系,以贵州省紫云苗族布依族自治县格凸河国家级风景名胜区中洞景区撂荒30余年的典型喀斯特次生林作为研究对象,基于1.68 hm²的固定样地,在实地调查的基础上,采用Hegyi竞争指数对样地内优势种群种内种间竞争关系进行定量分析。结果表明:1) 株数统计显示,共计调查胸径≥1 cm的林木2805株,其中亮叶桦、化香树、马尾松、枫香树和山杨共2455株,占总株数87.52%,为优势种群;2) 林木间的竞争压力主要来自其周围5m范围内的竞争木,除亮叶桦和枫香树的种间竞争大于种内竞争外,其余3个种群均为种内竞争大于种间竞争,表明次生林的竞争压力基本以种内竞争为主。5个优势种群种内竞争压力依次为化香树>山杨>马尾松>枫香树>亮叶桦;3) 竞争模型预测表明,优势种群的胸径与竞争强度之间呈现幂函数关系,揭示了随着林木个体胸径的逐渐增大,竞争强度减弱,当林木个体胸径在3~5cm时,竞争最为激烈,而当胸径达到10cm后,竞争强度的变化趋于平稳。因此,建议在林木胸径达到10cm之前,及时进行抚育管理,以减缓竞争压力,提高林分的稳定性。
关键词
林木竞争;喀斯特;优势种群;次生林;竞争指数
论文《喀斯特次生林优势种群竞争关系》发表在《林草资源研究》,版权归《林草资源研究》所有。本文来自网络平台,仅供参考。
林木竞争是指在森林生态系统中,林木间因争夺所在生境的资源(如光照和水分)而产生的互动关系,是林木对生存资源需求的体现,直接或间接地影响着林木的生长状况和未来长势。这种竞争不仅会扩大某些竞争木的生存范围、增加对邻近木的影响,而且还会影响群落结构和动态平衡,同时,也对群落的演替进程具有重要的影响。因此,林木间的竞争关系一直是森林生态与经营研究领域的热点之一。国内外学者利用多种数学模型对林木竞争强度进行了定量研究,以期更精准地预测林木生长,更科学地实施森林经营。Hegyi于1974年提出的与距离相关的单木竞争模型反映了个体对环境资源利用的能力大小,为个体间竞争的量化提供了有效的度量标准,对估计林木种内和种间的竞争强度具有良好的响应性,广泛用于分析林木间的竞争关系。
我国是世界上喀斯特面积最大的国家,以贵州省为中心的西南地区5.5×10⁵km²喀斯特分布区是世界三大喀斯特集中连续分布区之一。这种特殊的地貌,具有高度的空间异质性、复杂多变的小生境类型、群落结构单一和演替方向多样化等特征。由于喀斯特这一生态系统十分脆弱,加之频繁的人为干扰导致森林植被被破坏后恢复困难,在具有丰富物种多样性的同时,种内和种间竞争激烈,导致群落结构及生态系统不稳定。在喀斯特次生林中,林木竞争普遍存在,林木竞争的结果会导致参与竞争一方或者另一方生长衰弱甚至死亡。目前,尽管已有研究关注喀斯特地区单一树种的竞争关系,但对典型地带次生林群落中优势种群间竞争关系的研究还较缺乏。喀斯特地区次生林竞争关系的研究,有助于了解喀斯特次生林的发育和演替过程,预测喀斯特次生林的演替方向,为次生林植被恢复和质量提升提供理论依据。本研究以紫云苗族布依族自治县格凸河国家级风景名胜区中洞景区喀斯特典型次生林为研究对象,通过设置固定样地,利用Hegyi竞争指数定量分析5个优势种群种间及种内竞争关系特征,旨在揭示研究区次生林内不同优势种群的竞争关系,为合理调控森林结构,精准提升森林质量提供依据。
1 研究区与数据
1.1 研究区概况
贵州省紫云苗族布依族自治县(25°21′-26°3′N,105°55′-106°29′E),位于高原到丘陵过渡带的斜坡地带,属中、低谷盆地,属于亚热带季风性湿润气候,年均温15.3℃,年均降水量1337mm,气候垂直差异显著,四季分明。以紫云苗族布依族自治县具有鲜明喀斯特地貌特征的格凸河国家级风景名胜区中洞景区为研究区,该区普遍分布着非地带性的石灰土,土层浅薄,其成土母质主要由沉积岩和碳酸盐岩构成。研究区从20世纪80年代开始经历了从农耕到弃耕的转变,主要种植玉米的作物田逐渐荒废,演变为大面积的撂荒地带,经过30余年的自然演替和恢复,该区域目前人为干扰显著降低,表现出良好的自然生态恢复态势。
1.2 样地布设及人工调查
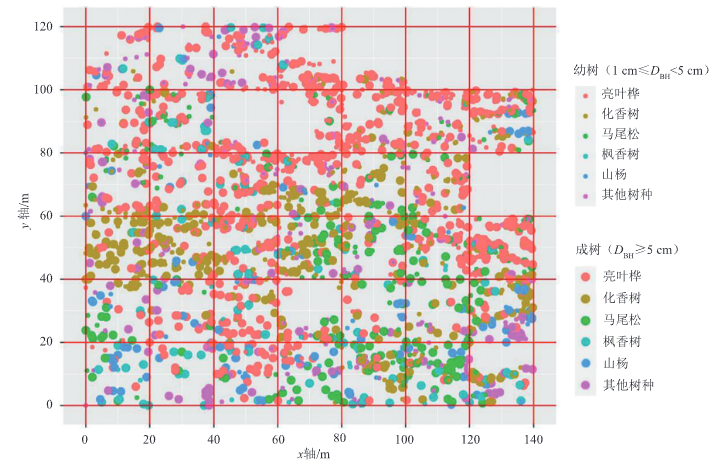
依据CTFS式(Centre for Tropical Forest Science)样地建设流程,选择具有代表性的地块,设置140m×120m的矩形样地,面积为1.68 hm²,并将大样地划分成42个小样地(20m×20m)。每木检尺的对象为样地内胸径≥1cm的林木,为每株林木钉上标识牌并记录唯一编号,以实现长期监测。测量其树高、胸径、枝下高、冠幅、生长状况等指标。同时,利用全站仪确定每株林木在样地中的相对坐标值(x, y)。
2 研究方法
2.1 径级划分
采用种群径级代替种群龄级的方法来研究喀斯特次生林优势种群结构。径级划分的方法参考池森等研究中的方法进行划分,以2cm为一个径级对喀斯特次生林内优势种群的胸径进行划分,共设置11个径级:Ⅰ径级(1~<3cm),Ⅱ径级(3~<5cm),Ⅲ径级(5~<7cm),Ⅳ径级(7~<9cm),Ⅴ径级(9~<11cm),Ⅵ径级(11~<13cm),Ⅶ径级(13~<15cm),Ⅷ径级(15~<17cm),Ⅸ径级(17~<19cm),Ⅹ径级(19~<21cm),Ⅺ径级(≥21cm)。其中亮叶桦(Betula luminifera)和枫香树(Liquidambar formosana)分别划分为11个径级,马尾松(Pinus massoniana)和化香树(Platycarya strobilacea)各划分为10个径级[样地中没有胸径≥21cm(Ⅺ径级)的林木],山杨(Populus davidiana)划为9个径级[样地中没有胸径≥19cm(Ⅹ径级、Ⅺ径级)的林木]。通常将1cm≤DBH<5cm的林木定义为幼树,DBH≥5cm林木定义为成树。
2.2 最佳邻域半径的确定
以样地内所有胸径≥1cm的林木作为对象木,以此为中心,以1m为步长,分别计算1~10m样圆半径内竞争木的竞争指数,以确定最佳竞争范围。随着样圆半径的增大,竞争指数逐渐减小,在1m样圆半径内,优势树种受到的竞争压力最大,而随着样圆半径的增大,优势树种受到的竞争压力逐渐减小。当样圆半径>5m时,竞争指数趋于平缓,因此,将最佳竞争范围确定为5m。
2.3 竞争指标的确定
采用Hegyi于1974年提出的单木竞争模型对5个优势种群的竞争指数进行计算,采用与距离有关的竞争指数作为竞争强度计算,其公式如下:
[I_{c}=sum_{i=1}^{N} I_{c, i}]
[I_{c, i}=sum_{j=1}^{n}left(D_{j} / D_{i}
ight) imesleft(1 / L_{i j}
ight)]
式中:(I_{C})为优势种群的竞争强度,其值越大,竞争越激烈;N为优势种群对象木株数;(I_{C, i})为第i株对象木的竞争指数;(D_{i})为对象木i的胸径(cm);(D_{j})为竞争木j的胸径(cm);(L_{i j})为对象木i与竞争木j之间的实际距离(m);n为第i株对象木周围的竞争木株数。
2.4 数据处理
利用Excel2016软件、R语言软件和Origin软件对数据进行统计与处理,使用forestSAS程序包计算Hegyi指数。
3 结果与分析
3.1 次生林树种组成及径级分布
调查到胸径≥1cm的木本植物2805株,其中亮叶桦、化香树、马尾松、枫香树、山杨的株数分别为1030、773、300、178、174株,共计2455株,占总株数的87.52%,将这5个树种作为优势种群。径级分布结果表明:5个优势种群径级分布不均匀,其中Ⅰ径级木数量较少,Ⅱ径级木数量最多,其次为Ⅲ~Ⅺ径级木,优势种群数量总体表现为幼树1081株(83.7%)小于成树1374株(90.7%),整体表现为种群更新不足。同时,Ⅰ径级木的数量较少,储备不足在一定程度上可能会影响群落的林分稳定性。优势种群的个体数量随着径级的增大而增加,到达Ⅱ径级后随着径级的增大其个体数量逐渐减少,5个优势种群的个体数量主要集中在Ⅱ~Ⅳ径级范围,共1931株,占总优势树种的78.66%。
3.2 次生林优势种群竞争指数比较
竞争指数结果显示,5个优势种群的总竞争指数排序为化香树(6311.87)>亮叶桦(5824.52)>马尾松(1628.07)>山杨(1008.93)>枫香树(897.83)。分别占总竞争指数的比例为40.28%、37.17%、10.39%、6.44%、5.73%。种间竞争指数排序为亮叶桦(3809.80)>化香树(2891.58)>马尾松(303.25)>枫香树(208.56)>山杨(199.98),分别占种间竞争指数的比例为51.39%、39.01%、4.09%、2.81%、2.70%。除亮叶桦外,各优势种群的种内竞争均较种间激烈,种内竞争顺序依次为化香树(3420.29)>亮叶桦(2014.72)>马尾松(1324.82)>山杨(808.95)>枫香树(689.27),分别占种内竞争指数的比例为41.42%、24.4%、16.04%、9.8%、8.35%。5个优势种群的种间竞争指数为7413.16,占总竞争指数的47.30%;种内竞争指数为8258.05,占总竞争指数的52.70%。整体表现为种内竞争大于种间竞争。
竞争指数均值结果表明,喀斯特次生林优势种群的竞争指数均值排序为化香树>山杨>枫香树>马尾松>亮叶桦。亮叶桦与化香树、马尾松、枫香树、山杨,以及化香树与马尾松存在显著性差异(P<0.01),其中化香树的竞争指数均值最大,亮叶桦最小。种间竞争指数均值从大到小顺序依次为枫香树>化香树>山杨>亮叶桦>马尾松。亮叶桦和马尾松、亮叶桦和枫香树、化香树和马尾松、化香树和枫香树、马尾松和枫香树、马尾松和山杨、枫香树和山杨存在显著差异(P<0.01),其中,枫香树最大,马尾松最小。种内竞争指数均值从大到小顺序依次为化香树>山杨>马尾松>枫香树>亮叶桦。亮叶桦和化香树、亮叶桦和马尾松、亮叶桦和枫香树、亮叶桦和山杨、化香树和马尾松、化香树和枫香树、化香树和山杨存在显著性差异(P<0.01)。其中,化香树最大,亮叶桦最小。
3.3 次生林优势种群不同径级竞争指数均值比较
优势种群不同径级竞争指数均值显示,亮叶桦和枫香树的种间竞争指数均值大于种内竞争,其余树种的竞争指数均值均为种内竞争大于种间竞争。5个优势种群竞争指数均值均在Ⅱ径级时最大,在该径级的竞争指数均值为439.40,占总竞争指数均值的52.00%。亮叶桦的竞争指数均值为300.54、化香树的竞争指数均值为311.09、马尾松的竞争指数均值为101.91、枫香树的竞争指数均值为65.01、山杨的竞争指数均值为66.29,其中,化香树的竞争指数均值最大,枫香树最小。亮叶桦、马尾松和山杨在Ⅱ径级以后,随着林木胸径的增大,竞争指数均值逐渐减小。化香树的竞争指数均值在Ⅱ~Ⅷ径级随着胸径的增大而减小,Ⅸ径级出现上升之后逐渐下降。枫香树的竞争指数均值在Ⅱ~Ⅴ径级随着胸径的增大而减小,Ⅵ径级出现上升之后逐渐下降。对比不同径级下5个优势树种的竞争指数均值进行比较发现,随着胸径的增大,Ⅰ~Ⅱ径级的竞争指数均值呈上升趋势,在Ⅱ径级达到最高值,之后逐渐下降并趋于稳定。
3.4 优势种群竞争指数与胸径模型构建
为分析胸径对竞争强度的影响,以优势种群胸径与其竞争指数进行回归分析,并利用指数函数、幂函数和线性函数3种回归模型进行对比分析。结果表明:指数函数对竞争指数的拟合效果不显著,线性函数评价指标不及幂函数评价指标,因此选择幂函数模拟和预测喀斯特优势种群种内和种间的竞争强度,其公式如下:
[I_{C}=A D_{BH}^{-B}]
式中:(D_{BH})是对象木胸径,A、B是模型系数,显著性检验结果均达到显著水平,模型的次幂为负值表示,随着对象木胸径的增大,竞争指数越小。
利用该模型模拟和预测喀斯特次生林优势种群的竞争强度。预测结果显示,优势种群的竞争指数随着胸径的增大而减小,当优势种群的胸径小于10cm时,其所受到的竞争压力最大,之后变缓;当优势种群的胸径大于10cm时,其所承受的来自种内和种间竞争的压力变化小,并保持在相对较低的状态。
4 讨论
本研究采用固定半径法来确定优势种群的最佳竞争范围。结果显示竞争指数随着样圆半径的增加逐渐减小,在样圆半径为5m时出现了拐点,5m之后竞争指数下降的幅度越来越小且逐渐趋于平缓。虽然随着样圆半径的增大,竞争木的数量在增多,但是竞争木对各对象木的竞争压力无明显下降的趋势,即在5m后竞争木对各对象木的影响微弱,因此,喀斯特次生林的优势种群受到的竞争压力主要来自其周围5m范围内的竞争木。申瀚文等对木荷(Schima superba),吴明对马尾松和麻栎(Quercus acutissima),柴宗政等对油松(Pinus tabulaeformis),许鲁东对香樟(Cinnamomum camphora)竞争关系的研究也表明了5m是林木竞争的最佳半径范围。Hou等通过构建考虑竞争影响的非线性混合效应模型来预测喀斯特地区多物种次生林中的冠幅宽度时,选择以5m作为固定半径来计算Hegyi竞争指数,结果显示这一选择显著提升了预测的准确性。
喀斯特次生林优势种群的种内竞争大于种间竞争,这一结论与王雁青等对喀斯特地区圆果化香树(Platycarya longipes)的研究结论基本相似,同一树种对资源的竞争通常比较激烈,是因为他们具有相同的生态习性与生态位,生态重叠范围大。此外,池森等的研究证明喀斯特地区的树种多为聚集分布,在小范围内同种林木数量较多,这也是研究区次生林种内竞争占主导的原因之一。虽然喀斯特次生林的竞争状况表现为种内大于种间竞争,但从不同树种的竞争现状来看,亮叶桦和枫香树的种间竞争大于种内竞争。这一研究结果与刘万生等对蒙古栎(Quercus mongolica),高浩杰等对红楠(Machilus thunbergii),刘虹佑等对泰山柳(Salix taishanensis)的种内种间竞争关系的研究结果相类似。分析其原因主要是喀斯特生境散布着裸露的岩石,其浅层土壤限制了林木生长的养分可用性,导致多数树种对生境的资源利用能力较低,不同的树种会为了生存而争夺资源,与其他树种展开竞争。亮叶桦作为群落的先锋树种,表现出较快的生长速度和优势地位,能够迅速郁闭成林,并在上层空间占据主导地位,其相应的种内竞争较小。次生林中阔叶树种类较多,数量庞大,生长习性多与亮叶桦相似,亮叶桦为维持其优势地位,势必会与其他树种发生激烈竞争。枫香树个体相对较小,竞争能力较弱,更容易受到来自其他树种的竞争压力。加之枫香树的种子主要是依靠风力传播,种子小且具翅,使得他们在高山多风的环境条件下散播具有很大的随机性,因此枫香树的种内竞争较小,种间竞争显著大于其他的种群,进一步表明了枫香树的他疏效应高于自疏效应。
对象木胸径的大小对林木竞争的影响较大。利用幂函数对对象木胸径与其受到的竞争强度进行拟合,结果表明,随着胸径的增大,喀斯特次生林优势种群受到的竞争强度逐渐减小,当5个优势种群胸径大于10cm时,其受到的竞争压力较小并逐渐趋于平缓。因此,在优势种群胸径大于10cm之前,应采取人工抚育管理来缓解竞争压力。喀斯特次生林优势种群结构特征及数量动态的研究表明,随着森林的演替,亮叶桦、马尾松和枫香树的数量优势将越来越明显。未来的森林群落可能会逐步向以亮叶桦、马尾松和枫香树为主的混交林演替。基于研究结果,考虑到优势树种的生存现状及竞争状态,有必要通过人为干预的手段来降低次生林竞争的激烈程度以促进其正向演替。以“去弱留壮、去密留疏、去弯留直”的原则进行采伐,主要采伐化香树和山杨等非目的树种。伐除部分大径级木,开辟林窗,减少主林层以及主林层对下层林木的竞争压力;保留长势良好的亮叶桦、马尾松和枫香树,以这些保留木为中心,对其周围竞争能力较强的树种进行抚育采伐;此外,采伐竞争力较强的Ⅱ径级林木以此来减少竞争木对有限的生存空间和资源的占用。由于样地内明显缺乏Ⅰ径级木,为提高目的树种幼苗幼树的存活率,防止林分更新不及时,可以考虑伐除周边影响其生长的杂灌,以改善生境质量。
5 结论
本研究采用Hegyi竞争指数模型对喀斯特次生林中亮叶桦、化香树、马尾松、枫香树和山杨5个优势树种的竞争关系进行调查与研究。结果表明,这5个种群共2455株,占总株数87.52%,为优势种群;研究区次生林的竞争压力主要来源于5m范围内的竞争木。5个优势种群,除亮叶桦和枫香树外,其余3个种群的种内竞争强于种间竞争,表明研究区次生林的竞争压力以种内竞争为主。5个优势种群的种内竞争强度排序为化香树>山杨>马尾松>枫香树>亮叶桦。优势种群的胸径与竞争强度符合幂函数关系,胸径增大时竞争强度逐渐减小。Ⅱ径级时(胸径3~5cm)竞争压力最大,当胸径达到10cm后,竞争强度的趋势平稳,因此,在优势种群胸径大于10cm前应实施人工抚育措施,调节喀斯特次生林的林分组成比例、优化林分结构、减轻竞争压力,从而维持喀斯特次生林的可持续发展。
参考文献
[1] 陶岩, 殷秀琴, 田育红, 等. 长白山红松针阔混交林种内、种间竞争关系研究[J]. 北京师范大学学报(自然科学版), 2011, 47(4): 428-431.
[2] 柳江, 洪伟, 吴承祯, 等. 天然更新檫木林竞争规律研究[J]. 江西农业大学学报, 2001(2): 240-243.
[3] 唐杨, 童跃伟, 韩艳刚, 等. 邻域竞争对长白山阔叶红松林关键树种生长的影响[J]. 应用生态学报, 2019, 30(5): 1479-1486.
[4] 许恒, 刘艳红. 珍稀濒危植物梓叶槭种群径级结构与种内种间竞争关系[J]. 西北植物学报, 2018, 38(6): 1160-1170.
[5] BELLA I E. A new competition model for individual trees[J]. Forest Science, 1971, 17(3): 364-372.
[6] LORIMER C G. Tests of age-independent competition indices for individual trees in natural hardwood stands[J]. Forest Ecology and Management, 1983, 6(4): 343-360.
[7] CHANDLER R, DEBELL D. Evaluation of field performance of poplar clones using selected competition indices[J]. New Forest, 2004, 27: 201-214.
[8] HEGYI F. A simulation model for managing jack-pine stands[C]// Fries J. Growth Models for Tree and Stand Simulation. Stockholm, Sweden: Royal College of Forestry, 1974: 74-90.
[9] 金则新, 朱小燕, 林恒琴. 浙江天台山甜槠种内与种间竞争研究[J]. 生态学杂志, 2004(2): 22-25.
[10] 李尤, 苏智先, 张素兰, 等. 珙桐群落种内与种间竞争研究[J]. 云南植物研究, 2006(6): 625-630.
[11] 黄伟程, 高露双, 赵冰倩. 不同间伐强度下竞争对东北阔叶红松林主要树种生长-气候关系的影响[J]. 北京林业大学学报, 2023, 45(1): 30-39.
[12] 汪清, 潘萍, 欧阳勋志, 等. 马尾松-木荷不同比例混交林种内和种间竞争强度[J]. 生态学杂志, 2021, 40(1): 49-57.
[13] FRAVER S, D’AMATO A W, BRADFORD J B, et al. Tree growth and competition in an Old-growth Picea Abies forest of Boreal Sweden: Influence of tree spatial patterning[J]. Journal of Vegetation Science, 2014, 25(2): 374-385.
[14] JI Yingrui, ZHANG Peng, SHEN Hailong. Competition intensity affects growing season nutrient dynamics in Korean pine trees and their microhabitats soil in mixed forest[J]. Forest Ecology and Management, 2023, 539: 539121018.
[15] 刘济明. 贵州茂兰喀斯特山地顶部森林群落种子库研究[J]. 林业科学研究, 2000(1): 47-53.
[16] WANG Linjiang, LUO Nana, SHI Qinglong, et al. Responses of soil labile organic carbon fractions and enzyme activities to long-term vegetation restorations in the Karst ecosystems, Southwest China[J]. Ecological Engineering, 2023, 194: 107034.
[17] 王琪, 容丽. 喀斯特石漠化区不同处理方式下滇柏的生长状况-以贵州省陈家寨为例[J]. 热带农业科学, 2016, 36(6): 37-44.
[18] FAN Zhouzhou, LU Shuyu, LIU Shuang, et al. The effects of vegetation restoration strategies and seasons on soil enzyme activities in the Karst landscapes of Yunnan, Southwest China[J]. Journal of forestry research, 2019, 31: 45-57.
[19] 杨文松, 容丽, 叶天木, 等. 黔中喀斯特次生林优势物种空间分布格局及关联性[J]. 应用生态学报, 2022, 33(5): 1215-1222.
[20] 蔡冰玲, 魏鑫, 范海兰, 等. 梅花山拟赤杨次生林优势树种种间竞争研究[J]. 西南林学院学报, 2009, 29(5): 16-18.
[21] 杨瑞, 喻理飞, 罗远飞, 等. 黔中喀斯特森林群落光皮桦种群竞争初步研究[J]. 贵州农业科学, 2010, 38(2): 153-155.
[22] 王雁青, 姚凯, 王晓双, 等. 茂兰喀斯特区圆果化香的种内和种间竞争强度分析[J]. 西北植物学报, 2021, 41(5): 880-888.
[23] 池森, 王从军, 黎庆菊, 等. 喀斯特次生林幼树更新空间分布格局及种间关联性[J]. 应用生态学报, 2020, 31(12): 3989-3996.
[24] 韦红艳, 徐铭, 柴宗政, 等. 喀斯特次生林优势种群结构及数量动态[J]. 东北林业大学学报, 2023, 51(7): 80-85.
[25] CONDIT R. Research in large, long-term tropical forest plots[J]. Trends in Ecology and Evolution, 1995, 10: 18-22.
[26] 陈科屹, 张会儒, 张博, 等. 长白山北坡天然次生林典型建群种的种群结构及动态特征[J]. 生态学报, 2021, 41(13): 5142-5152.
[27] 申瀚文, 鄢前飞, 曾思齐, 等. 木荷次生林种内和种间竞争研究[J]. 中南林业科技大学学报, 2012, 32(4): 81-85.
[28] 吴明. 马尾松-麻栎人工混交林种内与种间竞争关系研究[D]. 河南农业大学, 2016.
[29] 柴宗政, 王得祥, 张丽楠, 等. 秦岭西段油松天然次生林种群竞争关系研究[J]. 西北农林科技大学学报(自然科学版), 2012, 40(12): 57-63.
[30] 许鲁东. 香樟次生天然林群落空间分布格局及竞争关系研究[D]. 福建农林大学, 2022.
[31] HOU Rui, CHAI Zongzheng. Predicting crown width using nonlinear mixed-effects models accounting for competition in multi-species secondary forests[J]. PeerJ, 2022, 10: e13105.
[32] 姚慧芳, 卢杰, 王超, 等. 藏东南嘎朗国家湿地公园华山松种内与种间竞争强度研究[J]. 林业资源管理, 2021(3): 89-95.
[33] 刘万生, 李想, 陈福元, 等. 蒙古栎林种内和种间竞争研究[J]. 植物研究, 2020, 40(4): 552-558.
[34] 高浩杰, 高平仕, 王国明. 舟山群岛红楠林种内和种间竞争研究[J]. 植物研究, 2017, 37(3): 440-446.
[35] 刘虹佑, 马燕, 吴其超, 等. 濒危植物泰山柳种内与种间的竞争关系[J]. 森林与环境报, 2020, 40(2): 178-183.
[36] ZHANG Zhonghua, HU Gang, ZHU Jiedong, et al. Aggregated spatial distributions of species in a subtropical Karst forest, southwestern China[J]. Journal of Plant Ecology, 2013, 6(2): 131-140.
[37] 李冬林, 江浩, 王火, 等. 连云港市枫树湾景区枫香种群结构与分布格局[J]. 生态环境学报, 2020, 29(1): 49-58.
[38] 姚慧芳, 卢杰, 曾加芹, 等. 藏东南川滇高山栎天然林的种内与种间竞争指数的海拔差异[J]. 林业科学, 2022, 58(8): 53-62.
[39] 王丹, 王震洪. 枫香树种群格局与生境适应性的研究[J]. 山地农业生物学报, 2017, 36(3): 6-14.
[40] 李帅锋, 刘万德, 苏建荣, 等. 滇西北金沙江流域云南红豆杉群落种内与种间竞争[J]. 生态学杂志, 2013, 32(1): 33-38.



